![]()
![]()
Введение
Казалось бы, что Ростовская область, расположенная в степной зоне с открытыми безлесными пространствами, вряд ли может вызывать интерес разнообразием растительного покрова и богатством флоры. Это не Крым и не Кавказ! И все же!
Наличие крупных водоёмов (Азовское море, Цимлянское, Весёловское и Пролетарское водохранилища, озеро Маныч-Гудило), речных артерий (по территории области протекает около 150 больших и малых рек), песчаных массивов, меловых и каменистых обнажений, неоднородность почвенных и климатических условий, расчлененность рельефа делают растительный мир Ростовской области богатым и разнообразным.
На территории Ростовской области представлены такие типы растительности как степной, лесной, луговой, болотный, а флора области насчитывает более 2000 видов сосудистых растений [Федяева, 1994]. Кроме сосудистых растений во флоре области выявлено 148 видов мохообразных, 192 – лишайников, около 550 видов грибов – макромицетов и 648 видов фитопатогенных макро- и микромицетов [Федяева, Русанов, 2002].
Изучением растительности Ростовской области в конце ХIХ и начале XX веков занимались крупные ботаники – классики степеведения Г.Н. Высоцкий [1915], Б.А. Келлер [1907], И.К. Пачоский [1904, 1908, 1917], В.Н. Сукачёв [1902], К.М. Залесский [1918], а также В.Н. Баландин [1936], В.М. Богданов [1933], Г.И. Дохман и Т.И. Рыбакова [1933], Е.М. Лавренко [1924], А.Ф. Флёров [1932, 1935].
Пойменные луга в бассейне Дона изучали А.И. Бар [1919], Н.В. Властова [1919], С.Г. Горшкова [1919] и И.М. Крашенинников, опубликовавший совместно с С.Г. Горшковой и Н.А. Ивановой ряд работ [1922, 1928]. Растительности песков посвящены работы А.Г. Гаеля [1929, 1932], А.Д. Гожева [1929], А.Ф. Флёрова [1935].
Более двадцати лет посвятил изучению растительности Ростовской области профессор Ростовского госуниверситета И.В. Новопокровский [1915, 1921, 1931, 1933, 1940]. Им же была составлена мелкомасштабная карта растительности Ростовской области.
Во второй половине XX столетия были опубликованы работы А.П. Балаша [1955, 1959, 1961], большая часть которых отведена изучению Приазовских степей. Изучению лугов посвящены работы В.Н. Карпенко [1955, 1960], А.И. Луценко [1971, 1973].
Большой вклад в изучение естественных лесов степной части бассейна Дона внёс профессор Ростовского госуниверситета Г.М. Зозулин [1962, 1963, 1965, 1966, 1969, 1992].
Масштабные исследования растительности и естественных кормовых угодий Ростовской области проводились в течение 10 лет (1954-1964) коллективом геоботаников Ростовской землеустроительной экспедиции Института «Южгипрозем». Руководил исследованиями Б.Н. Горбачёв, разработавший метод растений-индикаторов, с помощью которого ему удалось составить крупномасштабную карту растительного покрова Ростовской области.
Метод основан на том, «что разные виды, входящие в состав коренного сообщества, неодинаково устойчивы к хозяйственным воздействиям. Одни из них быстро погибают при перегрузке пастбища скотом, другие выдерживают сбой и даже усиливаются под его влиянием. Некоторые не погибают даже при распашке и продолжают встречаться на пашне в качестве сорняков. Другие на пашне незаметны, но быстро восстанавливаются на залежах» [Горбачёв, 1974].
Изучение растительности и флоры Ростовской области продолжается и в настоящее время. В 1984-1985 гг. сотрудниками кафедры ботаники Ростовского госуниверситета была издана коллективная монография «Флора Нижнего Дона», а в 1996 году – справочник о редких, исчезающих и требующих охраны растениях, грибах и лишайниках. Общие сведения о современной флоре и растительности приводятся в обзоре, выполненном В.В. Федяевой [2002].
В 2004 году вышла в свет Красная книга Ростовской области, а с 2005 г. в рамках долгосрочного проекта «Ведение Красной книги Ростовской области» осуществляется мониторинг популяций редких видов.
Флора и растительность донских степей
Примечание: В тексте названия растений даны по-русски и по-латыни. Значками отмечены редкие виды, занесённые в Красную книгу Ростовской области (*) и в Красную книгу РФ (**). Приводятся самые распространённые синонимы названий растений как научные, так и народные (в том числе донских казаков). Латинское название и синонимы даются при первом упоминании вида. Таксономия и номенклатура соответствуют сводке С.К. Черепанова [1995], «Флоре европейской части СССР» [Т. I-VIII, 1974-1994], «Флоре Восточной Европы» [Т. IХ-ХI, 1996-2004].
Вся территория Ростовской области расположена в степной зоне. Степь, как тип растительности, представляет собой совокупность растительных сообществ засухоустойчивых и морозоустойчивых многолетних травянистых растений, среди которых первостепенная роль принадлежит дерновинным злакам.
Главным фактором образования степей является климат, отличающийся жарким, засушливым летом и морозной, часто бесснежной зимой. Сухость климата накладывает свой отпечаток на растительность и флору степи.
Летний зной, когда отсутствуют осадки, а температура воздуха достигает 40 градусов и выше, когда дуют сухие восточные ветры, понижая относительную влажность воздуха до 5 % (средняя – 62 %), без вреда для себя могут переносить лишь немногие растения, приспособленные к жизни в условиях длительной летней засухи и одновременно обладающие способностью переносить жёсткие условия зимы (глубокое промерзание почвы, ветры, зимние оттепели и т.п.). Это – растения сухотерпцы, или сухолюбы (по В.В. Алёхину), или ксерофиты [от греч. «ксерос» – сухой, «фитон» – растение].
«Роковая неподвижность растений лишает их возможности отойти в сторонку при воздействиях неблагоприятных факторов среды и вынуждает к решительным действиям по их нейтрализации или выключению» [Мордкович, 1982].
Ксерофиты в своём строении имеют целый ряд приспособлений (биологических, морфологических, анатомических, физиологических и др.), позволяющих им выживать в обстановке летнего зноя и отсутствия осадков. Способов приспособления к таким условиям выработалось много. Например, грыжник (грыжница) Бессера Herniaria besseri имеет сразу три способа приспособления к неблагоприятным климатическим условиям: низкий рост, мелкие листья и густое опушение. Ксерофиты легко отличить от растений других экологических групп (мезофитов, гигрофитов, гидрофитов) по целому ряду ксероморфных признаков:
1. Низкий рост и мелкие листья, часто рассечённые (что способствует уменьшению испаряющей поверхности) – тимьяны (чабрецы) Thymus, подмаренники Galium, живучка (дубровка) хиосская Ajuga chia и др.
2. Густое опушение из волосков, затрудняющих движение воздуха над устьицами (способствует уменьшению испарения) – шалфей эфиопский (ключ-трава) Salvia aethiopis, лютик иллирийский Ranunculus illyricus, остролодочник (остролодка, голубушка) волосистый Oxytropis pilosa, некоторые виды астрагалов Astragalus и др.
3. Сизый восковой налёт, отражающий солнечные лучи (предохраняет от нагрева и возможного ожога) – многие молочаи, например, молочай Сегье Euphorbia seguieriana, льнянка (по-местному – собачки) дроколистная Linaria genistifolia, вощанка (воскоцветник) малая Cerinthe minor и др.
4. Вертикальное расположение листьев – ребром к солнцу (спасает от перегрева) – хондрилла (хрящевница) ситниковидная Chondrilla juncea, степной сорняк – латук (салат) компасный Lactuca serriola, листья которого расположены не только вертикально, но и ориентированы с севера на юг (пример живого компаса).
5. Сворачивание листьев в трубку (при транспирации внутри её сохраняется влага, что уменьшает испарение) – многие степные злаки.
6. Редукция (исчезновение, недоразвитие) листьев (уменьшается испаряющая поверхность). Функцию фотосинтеза выполняют плотные, слабо испаряющие влагу стебли – спаржа (по-местному – холодок) лекарственная Asparagus officinalis, хвойник двухколосковый (эфедра, кузьмичёва трава, по-местному – степная малина) Ephedra distachya.
7. Выделение эфирных масел, за счёт летучести которых температура растения снижается.
«Пары эфирных масел часто горючи. Если поднести горящую спичку к такому растению в жаркий день, то масляные пары вокруг растения загорятся и будут гореть, пока не сгорят совсем. Самое же растение не загорается, потому что жидкое масло, содержащееся в нём, далеко не так горюче. На этом явлении, известном и в наших степях и в сухих частях Средиземноморской области, основано библейское предание о неопалимой купине: куст горел и не сгорал. Никакого чуда здесь, конечно, нет, и это явление относится к числу весьма обычных. Есть высокое травянистое южное растение ясенец (Dictamnus albus), с которым дочь знаменитого естествоиспытателя Линнея проделала опыт этого рода в Швеции, в Упсальском ботаническом саду, где оно разводилось [Буш, 1936].
Ясенец (бадьян), называемый старыми авторами – ясенец белый Dictamnus albus встречается и в нашей области в виде двух трудно различимых видов: ясенца кавказского Dictamnus caucasicus и ясенца голостолбикового Dictamnus gymnostylis. Чаще эти виды можно встретить в западных и юго-западных районах, реже – в Приазовье и северных районах. Места его обитания: каменистые степи, каменистые склоны балок, светлые байрачные леса и их опушки.
Внимание! При знакомстве с этим растением надо соблюдать осторожность. Растение ядовито и жгуче. При соприкосновении вызывает сильные ожоги кожи, которые проявляются через сутки в виде долго незаживающих волдырей, а после заживления на коже остаются тёмные пятна.
Большое количество эфироносов в семействе губоцветных: тимьяны, шалфеи Salvia, душица обыкновенная (материнка) Origanum vulgare и другие; а среди сложноцветных – полыни Artemisia. Вот чем объясняется неповторимый аромат степного воздуха и сена.
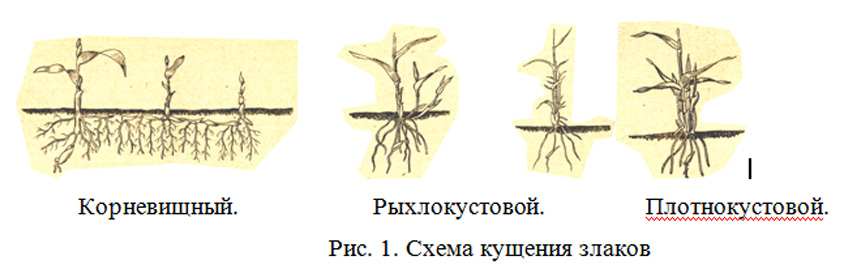
Особого внимания по своим приспособительным свойствам заслуживают злаки – типчак и ковыли – основные образователи растительных сообществ (ценозообразователи), строители (или эдификаторы) степных ценозов. Их листья узкие, часто волосовидные, свернутые в трубку, торчащие, из-за чего солнечные лучи на них падают косо, не нагревая листовую пластинку. Главная отличительная особенность степных злаков заключается в характере их ветвления, которое у злаков называется кущением.
В отличие от луговых злаков (рыхлокустовых и корневищных) степные злаки относятся к плотнокустовым или дерновинным. Их ветвление происходит в зоне кущения, расположенной у самой поверхности почвы. Новые побеги развиваются из боковых почек и растут внутри влагалища кроющего листа, не разрывая его. Таким образом, дочерние (боковые) побеги плотно прижаты к материнским и формируют компактный, плотный куст – дерновину. Число побегов в кусте плотнокустового злака – несколько сот, а иногда – свыше тысячи [Летняя практика, 1983]. С возрастом дерновина увеличивается, образуя кочку. Дерновина достигает в поперечнике 10 и более см. Она содержит много отмерших остатков старых стеблей и листьев, обладает свойством энергично впитывать и долго удерживать талую и дождевую воду.
Кочка степных злаков находится у поверхности почвы в чашевидном углублении. Это способствует стоку и задержке воды.
«Причина безраздельного господства злаков в травостоях степи заключается в их высокой приспособленности к сравнительно суровым и изменчивым условиям произрастания. Все злаки (особенно плотнокустовые) очень устойчивы к засухе, весьма умеренны к теплу и почвенному питанию, могут вегетировать практически на протяжении всего года, достаточно долговечны (дерновина типчака доживает до тридцати лет), а главное – располагают прекрасно отработанной системой вегетативного возобновления: отрастают за счёт почек, надёжно спрятанных от жары и зноя, от огня и травоядных животных внутри дерновин или ещё глубже – в самой почве» [Заповедник «Аскания-Нова», 1987].
Познакомимся с основными степными дерновинными злаками: ковылём, типчаком и тонконогом. Среди них выделяют мелкодерновинные – типчак и тонконог – и крупнодерновинные – все виды ковылей (за исключением ковыля Лессинга).
Ковыль Stipa. В злаковом травостое он занимает первое место. Этот род включает несколько видов, распространенных в Ростовской области. По морфологическому строению ковыли делят на перистые и неперистые – так называемые волосатики. У волосатиков – ость голая, гладкая, тонкая, как волос. У перистых – ость покрыта белыми волосками, пушистая, в виде пёрышка.
К волосовидным ковылям (в Ростовской области встречаются 2 вида) относится довольно распространённый ковыль волосовидный (волосистый, волосатик, тырса) Stipa capillata. Другой вид – ковыль сарептский (тырсик, тырсичка) Stipa sareptana встречается редко и только в крайних юго-восточных районах на солонцеватой почве.
Из перистых ковылей (в нашей области 8 видов) самый распространённый – ковыль Лессинга (по-местн. ковылок) Stipa lessingiana. Его легко отличить от других перистых ковылей по нескольким признакам. Это мелкодерновинный, мелкозёрный и раннецветущий ковыль. Его зерновка мелкая (0,6-1 см дл.), белёсая, волосистая по всей поверхности. Зерновки других перистых ковылей крупные (1,5-2,2 см дл.), желтовато окрашенные, почти голые (имеют лишь узкую полоску волосков и опушение внизу).
Ковыли различаются и по сезонному развитию. В течение вегетационного периода они сменяют друг друга (конечно, встречаясь и одновременно) в такой последовательности: первым начинает вегетацию и цветение ковыль Лессинга, за ним следует несколько ковылей – ковыль украинский Stipa ucrainica**, ковыль перистый Stipa pennata**, ковыль днепровский Stipa borysthenica*, за ними идут следующие ковыли – ковыль опушённолистный Stipa dasyphylla**, ковыль красивейший Stipa pulcherrima** и ковыль сарептский, затем следует ковыль узколистный Stipa tirsa*, а завершает это ковыльное буйство ковыль тырса, его массовое цветение начинается в середине июля и заканчивается в августе [Флора Нижнего Дона, 1985].
Если сгруппировать ковыли по их отношению к влаге и расположить по нарастанию ксерофильности (сухолюбию), то получится такой экологический ряд: ковыль узколистный – тырса – ковыль Лессинга – ковыль украинский – ковыль сарептский [Алёхин, 1951].
При определении ковылей можно также ориентироваться на места их нахождения и на степень встречаемости. Обычными, встречающимися во всех районах области являются ковыль Лессинга и тырса. Другие ковыли имеют меньшую степень встречаемости, иногда узкую локализацию.
После ковыля Лессинга и тырсы, чаще других в Ростовской области встречаются во всех районах: ковыль украинский и ковыль днепровский (последний предпочитает песчаные почвы в северных и западных районах). Реже встречаются: ковыль опушённолистный и ковыль красивейший (чаще в северных районах), ковыль перистый (чаще в северных и западных районах). Редко, в северных и западных районах – ковыль узколистный [Флора Нижнего Дона, 1985].
К наиболее распространенным степным дерновинным злакам относятся также типчак и тонконог.
Типчак (по-местн. типец) – один из видов рода овсяница Festuca. Его полное ботаническое название – овсяница валлисская Festuca valesiaca. Это мелкодерновинный злак с густой щёткой торчащих волосовидных, сизых (темно-зелёных с восковым налётом) листьев и с корнями в виде чёрной бороды. Соцветие – метёлка. По своей значимости фитоценотической значимости он занимает второе место после ковыля, а при повышении засушливости и засоления часто выходит на первое место. Он в большей степени, чем ковыль, устойчив к выпасу.
Тонконог (келерия ) – тонконог гребенчатый Koeleria cristata (по-местн. кипец). Это мелкодерновинный злак с колосовидной метёлкой (веточки метёлки тесно прижаты к оси, образуя плотное соцветие в виде «султана»). В злаковом травостое тонконог по обилию уступает ковылю и типчаку.
Среди степных растений есть и такие, листья и стебли которых не имеют ксероморфных признаков. Эти растения иным образом приспособились к перенесению засушливого периода. Познакомимся с ними.
Фреатофиты [от греч. «фреар» – колодец] – колодезные растения, по ним узнают, где копать колодец. Это растения, живущие за счёт влаги грунтовых вод. Они имеют мощно развитую стержневую корневую систему, в 10 и более раз превосходящую по массе надземные органы. Их корни уходят в почву на глубину 5-6 и более метров, достигая часто грунтовых вод. Их утолщённый стержневой корень играет с одной стороны роль мощного насоса для подъёма воды из больших глубин, а с другой - сам способен удерживать большое количество дефицитной влаги в своих тканях. Примером могут служить: верблюжья колючка обыкновенная (ложная, верблюдник, джантак, по-местн. янтак) Alhagi pseudalhagi, люцерна (по-местн. ямурка, медунка) румынская (степная) Medicago romanica, кермеки Limonium, астрагалы Astragalus.
Эфемероиды [от греч. «эфемерос» – скоро проходящий, быстро исчезающий]. Многолетние растения с коротким вегетационным периодом. Они развиваются весной, когда почва достаточно увлажнена, а к началу засухи уже успевают отцвести, отплодоносить, запасти питательные вещества и до следующей весны покоятся в почве в виде луковиц, корневищ, клубней.
Пример луковичных эфемероидов – гусиные луки Gagea, тюльпаны Tulipa, птицемлечники Ornithogalum, беллевалия сарматская (сарматский гиацинт) Bellevalia sarmatica**, корневищных эфемероидов – касатик карликовый (ирис, по-местн. петушок, кочеток) Iris pumila**.
Среди эфемероидов много красивоцветущих растений. Численность их сокращается из-за распашки целинных земель и массового бесконтрольного сбора. В связи с этим они быстро исчезают. Многие из них внесены в Красные книги РФ (**) и Ростовской области (*).
Эфемеры. Однолетние растения с короткой вегетацией. Они также как и эфемероиды используют весеннюю влагу, зацветают в марте, а в конце апреля уже плодоносят. В виде семян, защищённых от неблагоприятных условий семенной кожурой, они остаются в состоянии физиологического покоя до следующей весны. К ним относятся: веснянка весенняя Erophila verna, крупка перелесковая Draba nemorosa, рогоглфвник пряморогий (по-местн. репяшок) Ceratocephala testiculata, костенец зонтичный (зонтичник) Holosteum umbellatum и др.
Суккуленты [от лат. «succulentus» – сочный]. Это – растения с толстыми, мясистыми, сочными стеблями и листьями. Они обладают сильно развитой водоносной паренхимой с крупными водозапасающими клетками. Состав клеточного сока суккулентов таков, что позволяет им очень экономно расходовать воду. Недаром их называют «растениями-скопидомами» или «зелёными верблюдами». Примером могут служить виды растений семейства толстянковых: очиток едкий (расходник, ганец) Sedum acre, очитник пурпурный (очиток розовый Sedum telephium, заячья капуста) Hylotelephium trifillum.
Господствующий на открытых степных просторах ветер послужил причиной, вызвавшей приспособление растений к распространению плодов и семян с его помощью.
1. Хохолки. «У васильков и других сложноцветных растений степи мы находим хохолки из волосков на плодах. Благодаря этим хохолкам, действующим как парашюты, плоды долго держатся в воздухе и переносятся ветром на большие расстояния, содействуя распространению вида, захвату им всё большей и большей территории» [Буш, 1936].
2. Пушистые ости. Зерновки ковыля имеют пушистую ость, с помощью которой они не только удаляются от материнского растения, но и ввинчиваются в почву. Этому способствует и строение самой зерновки. Её нижний конец снабжён довольно длинным и тонким остриём, выше которого расположен «ёршик» из волосков, направленных кверху. Находящаяся у верхушки зерновки голая коленчато-изогнутая и спирально закрученная часть ости обладает гигроскопичностью. В сырую погоду ость раскручивается, удлиняется и загоняет зерновку в почву. При подсыхании ость скручивается, укорачивается, но зерновка, заякоренная ёршиком волосков, из почвы не вытаскивается.
Чтобы наблюдать за самозарыванием зрелой зерновки достаточно воткнуть её острым концом в одежду или в песок и смочить нижнее колено ости, как через несколько секунд ость начинает вращаться, а зерновка зарываться. Чтобы вытащить зерновку, надо приложить усилие, так прочно она удерживается волосками. Следует отметить, что зерновки ковыля, особенно тырсы, причиняют немалый вред овцам. Застревая в шерсти, а затем вбуравливаясь в кожу и мышцы, они вызывают тяжёлые заболевания и даже смерть.
3. «Перекати-поле». Некоторые травы при обильном ветвлении приобретают шаровидную форму. После созревания семян такой шар порывом ветра отламывается и, подгоняемый ветром, перекатывается по полю, рассеивая семена. Отсюда и название «перекати-поле». Такую форму образуют растения разных семейств: зопник (по-местн. железняк) колючий Phlomis pungens и шалфей эфиопский – сем. губоцветных; катран татарский Crambe tataria* – сем. крестоцветных; синеголовник полевой Eryngium campestre и резак обыкновенный Falcaria vulgaris – сем. зонтичных; качим метельчатый Gypsophilla paniculata – сем. гвоздичных; василёк раскидистый (верблюдка) Centaurea diffusa – сем. сложноцветных; углостебельник (гониолимон) татарский Goniolimon tataricum, кермек широколистный Limonium platyphyllum – сем. кермековых и некоторые другие.
Ошибочным является мнение о том, что растения «перекати-поле» лишь осенью в результате дождей и отгнивания нижней части стебля отделяются от корня и затем перекатываются. Перечисленные выше виды «перекати-поле» фенологически очень неоднородны. Они зацветают и заканчивают вегетацию в разное время. Так, например, когда катран татарский уже отрывается, зопник колючий и шалфей эфиопский – в полном цвету; качим метельчатый и гониолимон татарский зацветают, а резак и синеголовник ещё не зацвели. Таким образом, разные виды «перекати-поле» перекатываются в разное время, в том числе и летом. Но летом, на фоне зеленых и цветущих трав, они мало заметны. Главное предназначение формы «перекати-поле» – разбрасывание семян по степи. Вот здесь проявляется их особое приспособительное свойство. Оно заключается в постепенном рассеивании семян. При ударе о землю высыпаются не все семена, а лишь их часть. Эта способность рассеивать семена постепенно, равномерно и на большом расстоянии (что способствует захвату территории) обуслов
лена строением их плодов и семян.
Известный русский ботаник проф. В.В.Алёхин, изучавший особенности строения плодов «перекати-поля», задался вопросом: «Почему при перекатывании этих растений из сухих коробочек, перевернутых вниз, не высыпаются все семена сразу»? Оказалось, что у разных видов одного и того же рода, отличающихся образованием «перекати-поля», устройство плодов было разным. Так, у видов рода качим, не образующих «перекати-поле» (качим скученный, качим высочайший), коробочки в сухом состоянии открыты, их зубчики отогнуты наружу, и семена разбрасываются в стороны порывами ветра. У качима метельчатого – «перекати-поле» – зубцы коробочек всегда загнуты внутрь. Через узкую щель между зубцами семена высыпаются по одиночке и то только при сильном толчке.
У видов рода шалфей плоды (4 орешка) лежат на дне чашечки. Чашечки видов, не образующих «перекати-поле» (шалфей сухостепной, шалфей мутовчатый и др.), открыты вверх, и орешки свободно высыпаются при наклоне растения вниз. А вот у шалфея эфиопского – «перекати-поле» – зев чашечки густо оплетен массой волосков, образующих войлок, что сильно затрудняет высыпание орешков. Их выход из чашечки требует сильного толчка.
Таким образом, климат является мощным абиотическим фактором, влияющим на общий облик степных растений. О влиянии других абиотических факторов (эдафических, орографических) будет изложено далее.
Из группы биотических факторов особое значение имеет воздействие растений друг на друга.
Из всех типов растительности степной зоны степи обладают самым богатым флористическим составом. Они имеют очень высокую видовую насыщенность растительных сообществ (число видов, приходящихся на единицу площади). В северных степях (луговых, или разнотравных, распространенных в России севернее Ростовской области) на 1 кв. м приходится до 80 видов растений. В наших южных – настоящих, или ковыльных, степях – до 25 видов [Мордкович, 1982]. Такое обилие видов на небольшой площади возможно только благодаря сложившимся в течение длительного времени закономерностям пространственного размещения и связанного с ним фенологического развития растений.
Познакомимся с этими закономерностями.
1. Ярусное («поэтажное») распределение в пространстве как надземных, так и подземных частей растений. Важность этой закономерности отражена в оценке, данной В.В. Алёхиным: «Ярусность имеет глубокое экологическое значение, благодаря ей в сообществе могут произрастать растительные виды, являющиеся различными жизненными формами. Несомненно, здесь мы имеем результат очень длительного и сложного процесса взаимного приспособления растений друг к другу и к среде обитания. При этом подбирается вполне определённый комплекс видов, не только не мешающих друг другу, но имеющих преимущества в условиях совместного существования» [Алёхин, 1951].
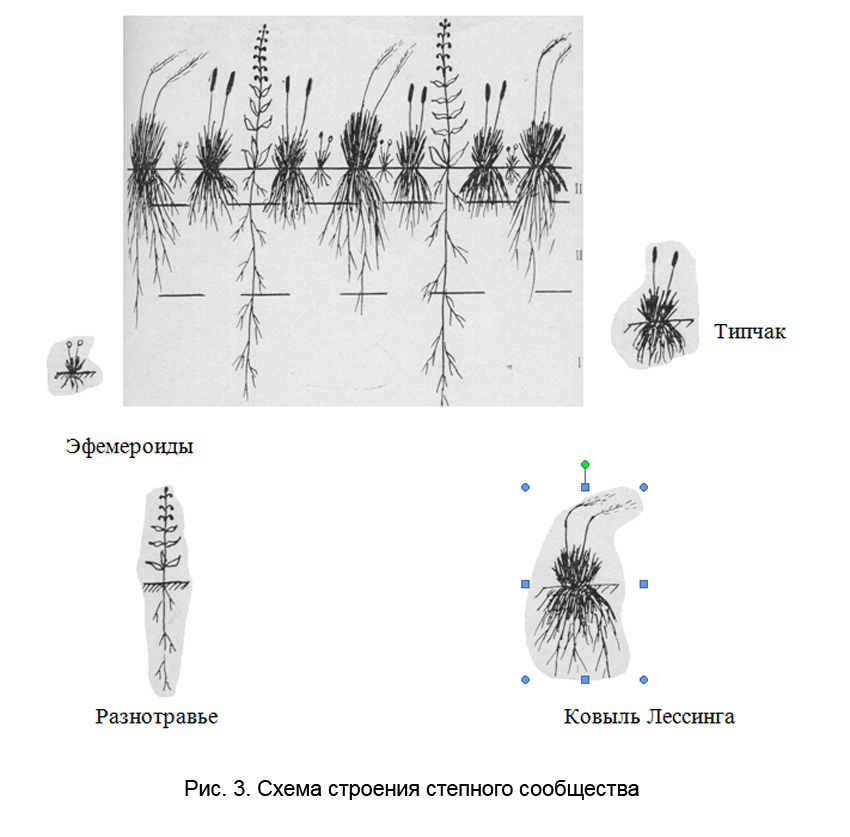
Дерновинные злаки, составляющие основу степного травостоя, имеют дерновины диаметром примерно в 10 см. Но их залегающая неглубоко, сильно разветвлённая мочковатая корневая система занимает значительно больше площади, чем дерновина. Корневые системы ковыля и типчака в почве смыкаются, а над поверхностью почвы их дерновины располагаются на некотором расстоянии друг от друга.
Промежутки между дерновинами злаков – кальвиции – заняты видами растений, составляющими разнотравье. Это в основном двудольные растения с длинными стержневыми корнями, проникающими через слои почвы, занятой корнями дерновинных злаков, и добывающими воду из глубоких слоёв, недоступных для корневой системы злаков. Ветвление у многих высокостебельных двудольных растений начинается не у самой поверхности почвы, и их свободные от ветвей нижние части стеблей легко размещаются между дерновинами злаков.
Среди степных злаков выделяют крупнодерновинные (ковыль) и мелкодерновинные (типчак и тонконог).
Рассмотрим надземные ярусы, образованные растениями разных жизненных форм. Отсчёт ярусов ведётся сверху.
Первый ярус составляют высокостебельные растения из красочного разнотравья (чаще это стержнекорневые двудольные). Поднимаясь выше злаков, они достаточно хорошо пропускают свет, необходимый для растений нижних ярусов. Классическим примером растения первого яруса может служить шалфей поникающий (по-местн. бабки) Salvia nutans. Его высокие безлистные стебли (листья собраны в розетку у основания стебля) выносят вверх поникающие соцветия и не затеняют растений нижних ярусов.
Второй ярус занят ковылём и видами разнотравья, имеющими среднюю высоту. Третий ярус составляют типчак и низкорослое разнотравье. В состав четвёртого яруса входят эфемеры, низкорослые эфемероиды и самые низкие, стелющиеся виды разнотравья. Можно отметить и пятый ярус, состоящий из мхов, лишайников и водорослей, расположенных непосредственно на поверхности почвы.
Существует ли закономерность в размещением подземных частей степных растений? Если надземная ярусность степного травостоя выражена менее чётко, чем в лесу (а в пустынной – полынно-дерновиннозлаковой степи она и совсем не выражена), то закономерность ярусного расположения подземных частей степных растений проявляется очень чётко (в том числе и в пустынной степи).
Вот как описывает подземную ярусность степного сообщества Б.Н. Горбачёв.
Имеются три основные яруса. Первый ярус составляют глубоко проникающие в почву стержневые корни многолетнего разнотравья, полукустарничков и кустарничков, второй – корни дерновинных злаков и третий – корни эфемеров и мелких эфемероидов. В формировании второго яруса, кроме дерновинных злаков, принимают участие также представители разнотравья, которые не имеют длинных стержневых корней и более крупные из эфемероидов (например, тюльпаны), иногда можно различить ещё четвёртый ярус из ризоидов мхов [Горбачёв, 1973].
2. Закономерность фенологического развития растений. Краски степного покрова богаты и непостоянны. В течение вегетационного периода степь неоднократно меняет свой облик. С ранней весны и до поздней осени через каждые 2-3 недели происходит смена внешнего вида – аспекта, который зависит от массового цветения тех или иных видов, господствующих в данный момент. Подобная смена фаз получилась в результате приспособления степных растений к различным отрезкам вегетационного периода. Это привело к тому, что каждый вид занял определённое место не только в пространстве, но и во времени.
Смена аспектов тесно связана с подземной ярусностью. Растения, корни которых развиваются в поверхностном горизонте почвы, вегетируют рано весной вследствие того, что эти горизонты прогреты раньше нижних и ещё не успели пересохнуть. Затем начинается аспект растений, корни которых составляют второй ярус и, наконец, аспект растений с глубоко проникающими стержневыми корнями. В степи в это время сменяются следующие аспекты [Горбачёв, 1973]:
Ранневесенний, когда цветут эфемеры и эфемероиды, а дерновинные злаки только начинают вегетировать. Степь нежно-зелёная с выделяющимися на её фоне яркими цветками тюльпанов и других крупных эфемероидов.
Поздневесенне-раннелетний, когда вегетируют и цветут типчак и ковыль, а вместе с ними и большинство представителей длительновегетирующего разнотравья. В это время степь очень красочна: сначала она густо зелёная от вегетирующих дерновинных злаков, а затем серебристо-белая от цветущего ковыля. На том и другом фоне выделяется пёстро цветущее разнотравье.
Позднелетний аспект, или аспект «выгорания». В это время дерновинные злаки и большая часть разнотравья уже закончили плодоношение, и вегетация их ослабляется, хотя полностью не прекращается. Степь становится жёлто-бурой или соломенно-жёлтой. Только в некоторых местах, где много ковыля тырсы, степь становится золотисто-зелёной, так как тырса, в отличие от остальных видов ковыля, цветёт в конце лета.
Раннеосенний аспект. В это время цветут поздние виды разнотравья и полукустарничков, имеющие длинный стержневой корень, главным образом, виды полыни. Степь принимает сизовато-серую, «седую» окраску.
Позднеосенний аспект. Под влиянием прошедших дождей и понижения температуры, степь снова зеленеет.
В степях встречаются рано цветущие (апрель-май) низкорослые (10-25 см высоты) осоки Carex, похожие на злаки.
Кроме травянистых растений в степи произрастают кустарники: миндаль низкий (миндаль степной, бобовник) Amygdalus nana, карагана кустарниковая (кустистая, дереза, чилижник, по-местн. сибирёк) Caragana frutex, роза (шиповник) собачья Rosa canina и роза майская Rosa majalis, боярышник (боярка, глог) обыкновенный Crataegus rhipidophylla, слива колючая (тёрн колючий) Prunus spinosa.
Степной травостой принято подразделять на три хозяйственно-биологические группы: злаки, бобовые и разнотравье. Степные злаки: ковыль, типчак, тонконог, а в более засушливых районах и житняк гребневидный Agropyron pectinatum. Из бобовых – люцерна румынская, астрагалы, остролодочник, эспарцет песчаный Onobrychis arenaria и др. Весь остальной травостой относится к разнотравью. Среди степного разнотравья встречаются представители многих семейств.
Степь – это не только растительный покров, это сложное природное сообщество – биогеоценоз, в котором взаимно связаны растения, животные, микроорганизмы, грибы, почвы.
Изучением почв Ростовской области занимался профессор Ростовского госуниверситета С.А. Захаров. Им, совместно с профессором И.В. Новопокровским, была разработана схема взаимодействия почв и растительности и их зависимость от изменения климата в Ростовской области.
Следует указать на планетарное значение степей, как единственного типа растительности, способного создавать плодороднейшие почвы планеты – чернозёмы. Великий почвовед В.В. Докучаев назвал чернозёмы «царём почв». Давая высокую оценку плодородия чернозёмов, он писал: «Чернозём для России дороже нефти, каменного угля, золотых и железных руд…Всё …ничто в сравнении с ним» [Гаврилюк, 1975].
Плодородие чернозёмов послужило причиной того, что большинство степных пространств было распахано и занято сельскохозяйственными культурами. Участки целинных степей сохранились лишь по неудобным для распашки склонам и балкам. Уцелели они на землях конезаводов, лесхозов, охотничьих хозяйств.
Небольшой экскурс в прошлое природы Ростовской области показывает, что целинные степи, когда-то занимавшие значительные площади, именовавшиеся Диким полем (так назывались степи между Доном и Днепром), стали усиленно распахиваться с 70-х годов ХVШ века. По мере роста населения интенсивность распашки возрастала. К середине ХIХ века степи занимали 60 % [Тараненко, 1975], а в настоящее время – лишь 17 % площади Ростовской области [Вальков, 2002]. Да и эти уцелевшие небольшие участки, подверженные интенсивной хозяйственной эксплуатации, сильно изменены.
Относительно нетронутых участков целины – этих крохотных и случайно уцелевших памятников природы, этих живых свидетелей величия наших степей сохранилось по всей области на особо охраняемых природных территориях не более 40 [Тараненко, 1975].
Об участках сохранившейся целинной степи в нашей области авторы книги «На разных широтах» в 1976 году писали: «В Ростовской обл. наибольший интерес представляют Персиановские степи между Новочеркасском и Шахтами. Первая Персиановская степь площадью 166 га относится к ксерофитному варианту приазовских степей. В ее флоре насчитывается 166 видов растений. Вторая Персиановская степь по характеру растительности сходна с предыдущей. Ее площадь 500 га, однако, в удовлетворительном состоянии находится только восточная часть».
Участки типчаково-ковыльной степи сохранились на берегах рек Юла и Маныч (Сальский р-н), на правом берегу р. Малая Куберле (Зимовниковский р-н) и по левому берегу р. Кундрючья (Белокалитвенский р-н).
Умеренно сухие типчаково-ковыльные степи с петрофитами (растениями скал и осыпей) и кальцефитами (растениями, предпочитающими богатые известью карбонатные почвы) расположены по левому берегу Северского Донца (Белокалитвенский р-н).
Приазовская разнотравно-типчаково-ковыльная степь сохранилась в балке Осиповской (Октябрьский р-н); она интересна тем, что здесь проходит северная граница ареала эремуруса представительного. По склонам балки Ясеневой и в верховьях балки Вяжа (Миллеровский р-н) можно наблюдать весь комплекс мезофильных и ксерофильных вариантов степей. В верховье р. Мечетки (Кашарский р-н) разнотравно-типчаково-ковыльная степь развита в комплексе с песчаной. Растительность развеянных песков в комплексе с песчано-ковыльной и песчано-типчаковой степью, а также осиновые колки можно видеть по левобережью р. Песковатки (Верхнедонской р-н).
На стыке Ростовской обл., Калмыкии и Ставрополь¬ского края расположено оз. Маныч-Гудило, в долине которого и на его многочисленных островах, можно наблюдать разные варианты долинных степей и галофильную (солеустойчивую) растительность. На озере можно полюбоваться и различными редкими птицами – лебедями, пеликанами и др. [Белоусова Л.С., Денисова Л.В., 1976]. С 1995 г. здесь существует Государственный природный заповедник «Ростовский». Площадь заповедника составляет около 9,5 тыс. га, а его охранной зоны – более 74 тыс. га.
О трагической судьбе степей В.Г. Мордкович [1982] писал так: «Если будет заведена «Красная книга экосистем», то степь в неё занесут в первую очередь. Среди всех экосистем мира судьба степей наиболее драматична. Главным героем последних актов этой драмы является человек. История цивилизации так тесно и причудливо сплетена с жизнью степных экосистем, что человечество просто обязано от щедрот своих пожертвовать на сохранение этого вымирающего ландшафта… На что может рассчитывать человечество? Как скоро мы можем получить результат своих усилий по восстановлению природных экосистем? С какой скоростью идёт процесс восстановления естественным путём? Скорость этого процесса можно проиллюстрировать хотя бы примером с почвами. Установлено, что для образования слоя чернозёмной почвы толщиной в 1 см требуется 50-100 лет, т.е. наше богатство – чернозёмные почвы – естественным путём формировались минимум 3-5 тысяч лет».
Чтобы степи не исчезли бесследно с лица Земли, а вместе с ними не исчезли и чернозёмы, степи надо охранять! Но как? По этому вопросу существует несколько точек зрения. Рассмотрим некоторые из них.
Один литературный источник [Травянистые растения СССР, 1971] рекомендует самую строгую охрану. Другой же [Вальтер, 1975] предупреждает «При полном заповедовании степей происходит деградация растительного покрова, поскольку интенсивно накапливающаяся подстилка угнетает дерновинные злаки. Мёртвый покров снижает испарение, благодаря этому улучшается водный режим почвы, что в свою очередь способствует поселению корневищных злаков – пырея (Elytrigia repens), вейника наземного (Calamagrostis epigeios)». Изменяются почвенные процессы, ухудшается прогревание верхних слоёв почвы. Не способные конкурировать в этих условиях ковыли и типчак изреживаются и выпадают. На освободившихся местах поселяются более мезофильные виды растений, несвойственные целинной степи. Происходит мезофитизация степной растительности, известная под названием «олуговение степи». Так случилось в 60-е годы прошлого столетия с памятником природы «Персиановская степь».
Как указывает Б.Н. Горбачёв, деградация степи при строгой охране может идти и по пути «забурьянивания», когда на освободившихся от дерновинных злаков местах поселяются бодяки Cirsium, чертополохи Carduu, татарник колючий Onopordum acanthium, т.е. глубоко укореняющиеся сорняки, которые вытесняют типичные степные растения. Степь превращается в бурьянник, похожий на залежь [Горбачёв, 1973].
В естественных условиях, до заселения степей человеком (см. примечание) на широких степных просторах нашей области паслись тарпаны (дикие лошади), сайги или сайгаки (сородичи настоящей антилопы). В результате пастьбы удалялась лишняя листовая масса, а перемещение по степи копытных животных способствовало разбиванию кочек дерновинных злаков, что в свою очередь приводило к удалению старого сухого травостоя (ветоши, старики, колдана, или колтуна). Только при таких условиях поддерживается равновесие степной экосистемы.
Примечание: «По свидетельствам археологов, люди появились на территории теперешней Ростовской области 350-100 тысяч лет назад. Они оставили после себя каменные орудия, найденные на стоянках в низовьях Северского Донца и на побережье Таганрогского залива. Люди того времени были собирателями и охотниками. Земледелие на Дону зародилось около 4-х тысяч лет назад. Распашка же целины по-настоящему началась здесь лишь во второй половине XVIII века» [Тараненко, 1975].
Как же создать такие условия? Могут ли роль диких копытных животных выполнять наши домашние копытные? Да, могут! Но только при умеренном выпасе. Поедание части растительной массы не только безвредно, но даже необходимо (как это видно из ранее приведённого примера). Кроме того, животные втаптывают семена растений в почву, улучшают условия их прорастания, а, перенося семена на копытах и шерсти, способствуют их расселению.
Какая судьба ждёт степь при чрезмерной пастбищной нагрузке? При чрезмерной нагрузке, при перевыпасе, степное сообщество деградирует, постепенно разрушается и исчезает. Скот уплотняет почву, способствуя её иссушению, вытаптывает растения копытами, что особенно вредно для дерновинных злаков, а, поедая в первую очередь более ценные в кормовом отношении ковыль и типчак, он полностью лишает степь её основных образователей.
Это явление носит название сбоя, или пасквальной дигрессии [от лат. «pascuum» – пастбище и «digredior» – удаляться]. Если сбой продолжается, то сначала остаются непоедаемые растения: полыни Artemisia, дурнишники Xanthium, рогач песчаный (по-местн. серко, устели-поле, утри-хвост, эбелек) Сeratocarpus arenarius и др. Сильно разрастается мятлик живородящий Poa crispa. При дальнейшем сбое исчезают и эти растения. Сообщество полностью погибает, остаётся голая земля.
Вывод: для поддержания типичной степной растительности необходим умеренный выпас, каким он был в доисторическое время [Вальтер, 1975].
Степь Ростовской области неоднородна. На её характер влияет засушливость климата, обусловленная количеством выпадающих осадков и испаряемостью, которая, в свою очередь, зависит от температуры воздуха и силы ветра. В нашей области засушливость усиливается в юго-восточном направлении, в этом же направлении меняется и характер степи.
По мере ксерофитизации степь теряет свою красочность: в ней исчезают виды разнотравья (как наиболее влаголюбивые, мезофильные), остаются злаки и появляются полыни (как наиболее засухоустойчивые, ксерофильные), травостой становится низким и разреженным, всё больше появляется пятен голой земли. Чернозёмы (на севере, западе и юго-западе) сменяются тёмно-каштановыми, каштановыми и светло-каштановыми часто засолёнными почвами (на востоке и юго-востоке).
По показателям, отличающим один тип степи от другого (видовая насыщенность, состав образователей, экологическая структура разнотравья, сезонное развитие) в нашей области выделяются, сменяющие друг друга с запада на восток, три степных подзоны, и, соответственно, три подзональных типа степной растительности: настоящие (разнотравно-дерновиннозлаковые), сухие (дерновиннозлаковые) и пустынные (полынно-дерновиннозлаковые) степи.
Современное состояние степей (почти повсеместная распашка и усиленное хозяйственное использование сохранившихся степных целин) затрудняет установление точных геоботанических границ между названными подзональными типами.
Сохранившиеся, небольшие по площади степные участки представляют большую научную и практическую ценность. Степи важны, прежде всего, как эталоны высокопродуктивных растительных сообществ, постоянно повышающих плодородие земель. Именно степные сообщества формируют чернозёмы.
Степи являются резерватами очень многих хозяйственно ценных видов растений, в числе которых пищевые, лекарственные, медоносные, технические, эфирно-масличные, кормовые и другие полезные растения.
Особого внимания заслуживают редкие и исчезающие виды, число которых возрастает в связи с сокращением площадей целинных земель и возрастанием антропогенной нагрузки на сохранившиеся степные участки.
В степных сообществах находят убежище многие виды позвоночных и беспозвоночных животных, немалая часть которых находится на грани исчезновения.
Таким образом, степи имеют огромное биогеоценотическое, природоохранное, экономическое и эстетическое значение в планетарном масштабе.
Настоящая степь
Настоящая степь приурочена к наименее засушливым районам области, занимает наиболее высокие плоские водоразделы в районах Доно-Донецкой возвышенности и Донецкого кряжа, а также водораздельные пространства в западной части Ейско-Егорлыкской равнины [Горбачёв, 1974].
Этой степи в прошлом принадлежала основная часть территории области. Она занимала северную, западную и южную части, гранича на востоке с Обливским, Морозовским, Орловским районами, а на юге – с Краснодарским краем. В полосе своего распространения настоящая степь неодинакова. Можно различить северную и южную её части. Северная часть занимает районы Доно-Донецкой возвышенности, а южная – районы Ейско-Егорлыкской равнины. Южную часть настоящей степи И.В. Новопокровский [1940] относил к особому Приазовскому району.
Сформированная на чернозёмах настоящая степь больше других степей пострадала от распашки. В настоящее время она занимает 569,6 тыс. га, что составляет лишь 5,64 % территории Ростовской области, площадь которой 10096,7 тыс. га. Если сохранившиеся участки трёх подзональных типов степей принять за 100 %, то на настоящую степь приходится 32,6 % [Федяева, 2002].
Каковы же характерные особенности настоящей степи? Что её отличает от сухой и пустынной степей? В самом названии – разнотравно-типчаково-ковыльная – заложено основное её отличие. Она богата разнотравьем, делающим её красочной и живописной.
Настоящая степь имеет следующие отличительные признаки:
1. В травостое значительное участие принимает красивоцветущее и пёстрое разнотравье.
2. Кроме основных плотнокустовых, узколистных (ксерофильных) злаков (типчака, ковыля, тонконога), в растительном покрове участвуют рыхлокустовые и корневищные, широколистные (мезофильные) злаки.
3. Высокий и густой травостой, что обусловливает проективное покрытие от 70 до 100 % и высокую продуктивность (выход сена с единицы площади), которая в зависимости от интенсивности хозяйственной эксплуатации колеблется от 100 до 350 г с 1 кв. м [Федяева, 2002].

(разнотравно-дерновиннозлаковая, или разнотравно-типчаково-ковыльная)